ЛЕОНАРД
ВЛАДИМИРОВИЧ ПОЛИЩУК
|
|
Профессор кафедры общей экологии биологического факультета МГУ имени М.В. Ломоносова, доктор биологических наук Контактные адреса: 119992,
Россия, Москва, Ленинские горы, д. 1, стр.
12, Биологический факультет МГУ, к.216 Телефон: +7(495)939-38-72 e-mail: leonard_polishchuk@hotmail.com |

_____________________________________________________________________________
Доклады на семинаре "Популяционная и системная экология"
Научные интересы
Если попытаться в двух словах определить область моих научных интересов, то
я бы назвал ее демографическим анализом. Демографический – потому что меня
всегда интересовали и продолжают интересовать темп размножения (рождаемости) и
темп гибели (смертности) организмов, т.е. демография животных. Анализ – потому
что рождаемость и смертность выступают как непосредственные причины изменения
численности, тогда как факторы внешней среды не могут воздействовать на
численность иначе чем через посредство рождаемости и смертности. Это
промежуточное, а на самом деле центральное положение рождаемости и смертности в
комплексе факторов, определяющих характер изменения численности, позволяет,
опираясь на рождаемость и смертность, реконструировать последовательность
причин и следствий, ведущих от факторов внешней среды к изменению численности
популяции. В более широком плане демографический анализ связан с изучением не
только динамики численности, но и динамики биомассы (которая, наряду с
рождаемостью и смертностью определяется скоростью роста особей и темпом
продуцирования популяции) и соответственно всего спектра биологических
показателей, влияющих на рождаемость, смертность, скорость продуцирования и
скорость элиминации биомассы. Только что сказанное относится в основном к
локально-популяционному аспекту демографического анализа. Но демографические
характеристики можно рассматривать и в сравнительно-видовом аспекте, т.е.
оперировать средними, характерными для вида показателями (такими как средний
размер помета или средняя продолжительность жизни после достижения половой
зрелости). В этом случае демографические характеристики по-прежнему занимают
промежуточное положение в цепи фактор-численность, но служат чаще не для
объяснения динамики численности, а для прогноза среднего уровня численности
(плотности населения) вида по сравнению с другими видами или же, например, для
прогноза вероятности вымирания. В последние годы такой сравнительно-видовой
подход стали называть макроэкологией. Таким образом, демографический анализ
позволяет перекинуть мостик от динамики численности данной локальной популяции
(«микроэкологии») к макроэкологии. Динамикой численности и биомассы я занимался
в основном на планктонных ракообразных, в частности дафниях, а
сравнительно-видовыми зависимостями – на млекопитающих. Такой выбор объектов
определяется моим «бэкграундом» (я заканчивал кафедру зоологии беспозвоночных,
где моим научным руководителем был проф. А.М. Гиляров) и, конечно, задачами
исследования (дафнии в силу своих малых размеров и высокой скорости размножения
особенно удобны для изучения динамики численности, а млекопитающие в силу
высокой изученности этой группы – для сравнительно-видовых построений).
Удивительно здесь то, что хотя всякая задача решается на конкретных организмах и
вообще в конкретных, частных обстоятельствах, ответы на частные вопросы могут
иметь общее значение. Почему так получается – не совсем ясно. Отчасти,
возможно, потому, что частный вопрос с самого начала удается поместить в общий
контекст, каковым, например, может быть теория жизненных стратегий или теория
вымирания (при всей слабости имеющихся в настоящее время экологических теорий).
А отчасти потому, что исследователи ищут – и, если повезет, находят – более или
менее общие методы, инструментарий или математический аппарат, пригодный для
решения многих частных проблем. Общие же методы часто приводят к общим
результатам.
В последние годы моя работа
развивалась в основном по трем направлениям:
1. Динамика массы тела и стратегия распределения ресурсов при изменении
пищевых условий (на примере ветвистоусого рачка Daphnia galeata)
Общий метод, примененный в этой работе, – анализ вкладов (contribution analysis). Суть метода состоит в том, что, во-первых, мы выделяем некоторый показатель – здесь это масса тела Ct, во-вторых, строим зависимость (модель) этого показателя от более элементарных и вместе с тем информативных экологических характеристик – здесь это стандартная (приведенная к фиксированному линейному размеру особей) соматическая масса D, число яиц в кладке Ea и средняя масса одного яйца Ce, в-третьих, выбираем траекторию движения системы в пространстве (D, Ea, Ce) (эта траектория определяется испытанными концентрациями пищи – каждой концентрации соответствует своя тройка (D, Ea, Ce), а концентрации, в свою очередь, выбраны исходя из пороговой и начальной лимитирующей концентрации для D. galeata), в-четвертых, используя аппарат частных производных, вычисляем, как затраты на увеличение стандартной соматической массы (образующие вклад Con D), числа яиц в кладке (вклад Con Ea) и массы одного яйца (вклад Con Ce) влияют на изменение общей массы тела ∆Ct при увеличении концентрации пищи (вклады выражаются в тех же единицах, что и масса тела, причем Con D + Con Ea + Con Ce = ∆Ct). Показано, что в области низких значений концентрации (при увеличении концентрации от 0.07 до 0.11 мг С л‑1) дафнии в основном увеличивают соматическую массу и гораздо меньше ресурсов вкладывают в увеличение числа яиц и массы яйца; при дальнейшем увеличении концентрации (от 0.11 до 0.18 мг С л‑1) примерно равное количество ресурсов затрачивается на увеличение соматической массы и числа яиц и гораздо меньшее – на увеличение массы яйца; наконец, когда концентрация намного превосходит начальную лимитирующую (при ее увеличении от 0.18 до 1.00 мг С л‑1), т.е. пища перестает быть лимитирующим фактором, основное количество ресурсов расходуется на увеличение числа яиц, а масса яйца даже уменьшается (рисунок). Эти результаты находят свое объяснение в свете теории жизненных стратегий. Сдвиг приоритетов в распределении ресурсов при улучшении пищевых условий интерпретируется как механизм, обеспечивающий первоочередное выживание взрослых особей в условиях недостатка пищи и высокую скорость размножения – в условиях избытка.
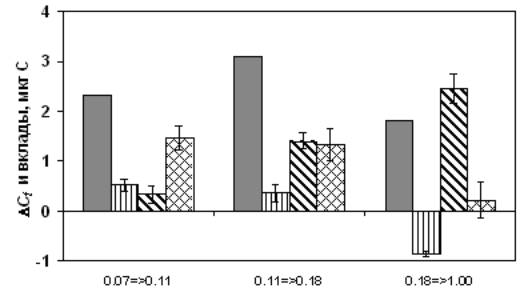
![]()
2. Демографические характеристики и вероятность оказаться под угрозой вымирания (на примере млекопитающих)
Современная теория вымирания говорит, что, во-первых, вымирание носит
вероятностный характер и, во-вторых, вероятность вымирания зависит от скорости
роста численности: чем выше эта скорость, тем ниже вероятность вымирания вида.
Согласно классическому (частотному) определению вероятности, для оценки
вероятности вымирания необходима однородная совокупность видов со сходными
демографическими характеристиками (а значит, со сходной скоростью роста
численности), одни из которых вымерли, а другие – нет. Тогда доля вымерших
видов от общего числа видов будет искомой вероятностью при данной скорости
роста численности. Очевидно, что в современной фауне таких совокупностей просто
нет (хотя их можно попытаться найти на палеонтологическом материале). Зато
современная фауна включает как виды, находящиеся под угрозой вымирания (их
заносят в Красную книгу; среди млекопитающих, например, таких видов 25%), так и
виды, находящиеся в настоящее время в безопасности («некраснокнижные»). Этот факт
открывает путь к оценке вероятности оказаться под угрозой вымирания. На
основании анализа более 130 литературных источников был собран материал по
демографии и массе тела 90 видов млекопитающих из фауны бывшего Советского
Союза, из которых 25 видов занесены в «Красную книгу СССР» (2 изд., 1984). В
выборку были включены все виды, для которых удалось найти необходимые данные.
По этим данным для каждого вида были рассчитаны следующие демографические
показатели: плодовитость за год и за время жизни, размер помета и
продолжительность жизни взрослых особей, и все 90 видов были ранжированы по
каждому показателю, а также по массе тела. Даже беглый просмотр ранжированных
видов показал, что, как и предсказывает теория вымирания, в Красной книге
представлены в основном крупные, медленно размножающиеся виды. Для того, чтобы
найти вероятность оказаться под угрозой вымирания и построить количественную
зависимость между этой вероятностью, с одной стороны, и демографическими
показателями и массой тела, с другой, был использован аппарат логистической
регрессии. Эта регрессия чрезвычайно удобна для решения поставленной задачи,
поскольку она специально ориентирована на построение зависимости двоичного
признака (здесь – присутствие / отсутствие вида в Красной книге) от обычного
количественного и, кроме того, интерпретируется как вероятность (в частности,
потому что она заключена между 0 и 1). Среди пяти изученных признаков наиболее
тесно связанной с интересующей нас вероятностью оказалась годовая плодовитость
(рисунок; зависимость, представленная на рисунке, названа функцией уязвимости).
Все краснокнижные виды в нашей выборке, среди которых такие хорошо известные
как зубр, тигр и русская выхухоль (рисунки зверей взяты с сайта http://www.nature.ok.ru), имеют относительно
низкую плодовитость, тогда как большинство некраснокнижных видов находятся на
противоположном участке диапазона плодовитости (в качестве примера показан
кролик с его высокой, вошедшей в поговорку скоростью размножения; рисунок с
сайта http://www.animalls.net). Полученные оценки вероятности оказаться под
угрозой вымирания предлагается использовать для определения природоохранных приоритетов,
в частности для разумного распределения усилий при мониторинге видов с разной
годовой плодовитостью (конечно, это предложение не исключает, а дополняет
использование других критериев). Так, основного внимания заслуживают виды, для
которых эта вероятность превышает некоторую малую величину. В качестве такой
величины предлагается взять 0.01; ей соответствует показанная на рисунке
пороговая годовая плодовитость Bcr = 2.9 дочерей/год. Фактически в
нашей выборке нет ни одного краснокнижного вида среди 44 видов с плодовитостью
больше 1.75 дочерей/год (последняя величина характеризует выхухоль; рисунок).
Таким образом, 2.9 дочерей в год – пороговая плодовитость с хорошим «запасом
прочности».
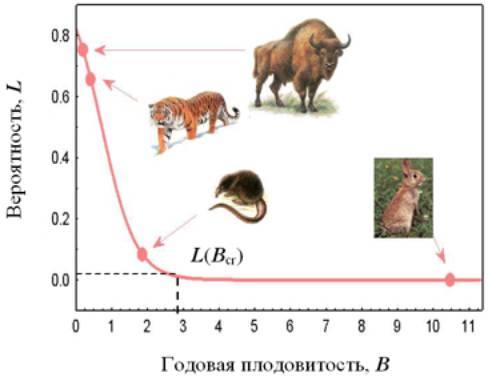
3. Динамика рождаемости и соотношение контроля со стороны нижнего и верхнего трофических уровней (на примере ветвистоусых ракообразных, главным образом дафний)
Общий метод, примененный в этой работе, – тот же, что и в работе по
динамике массы тела, – анализ вкладов. Только здесь в качестве основного
показателя взята удельная рождаемость b (рождаемость в расчете на одну
особь), а в качестве более элементарных характеристик, определяющих
рождаемость, – плодовитость F, долявзрослых особей A и скорость развития яиц V = 1/De, где De – продолжительность
развития яиц. Между этими характеристиками и факторами внешней среды имеется
определенное соответствие: плодовитость, как правило, зависит от пищевых
условий, доля взрослых особей в популяции часто (хотя и не всегда) находится
под контролем размерно-избирательной элиминации со стороны хищников –
беспозвоночных, потребляющих в основном молодь, или рыб, потребляющих в
основном взрослых дафний, а скорость развития яиц определяется почти
исключительно температурой. Удельная рождаемость связана с указанными
характеристиками с помощью хорошо известной в планктонологии формулы (модели)
Палохеймо: b = V ln(1 + F A). Траектория движения системы определяется в данном случае
временнóй динамикой рождаемости и определяющих ее характеристик, а
вклады представляют собой компоненты изменения рождаемости, которые зависят от
изменения соответствующих характеристик и в сумме дают изменение рождаемости на
данном временном интервале, т.е. ∆b = Con F + Con A + Con V. Имеющиеся материалы
показывают, что в водоемах, где хищников нет, ведущим фактором динамики
рождаемости является плодовитость (т.е. |Con F| > |Con A|), а там, где велик пресс хищников, – доля
взрослых (т.е. |Con F| < |Con A|). В водоемах умеренного климата в течение
большей части вегетационного сезона скорость развития яиц не является ведущим
фактором динамики рождаемости на интервалах времени порядка De (т.е. |Con De| < |Con F| и |Con De| < |Con A|). По-видимому, то или
иное соотношение абсолютных величин вкладов плодовитости и доли взрослых в динамику
рождаемости, |Con F|/|Con A|, может служить индикатором того, контролируется
ли популяция пищей, т.е. со стороны нижнего трофического уровня (bottom-up effect), или хищниками, т.е. со стороны верхнего трофического уровня (top-down effect). Относительная роль контроля «снизу» и «сверху» является одной из
наиболее горячо обсуждаемых в экологии проблем.
Ключевые слова: демография, популяционная экология, динамика численности, рождаемость,
смертность, плодовитость, масса тела, стратегия распределения ресурсов в
меняющихся условиях среды, контроль изучаемой популяции со стороны нижнего и
верхнего трофических уровней, анализ вкладов (contribution analysis), Daphnia, макроэкология, вероятность оказаться под
угрозой вымирания, Красная книга, логистическая регрессия, млекопитающие
Учебно-педагогическая деятельность
В течение примерно 10 лет мною читался курс «Введение в количественную
экологию» на кафедре зоологии беспозвоночных Биологического факультета МГУ
(совместно с А.В. Смуровым), на основе которого было написано учебное пособие
«Количественные методы оценки основных популяционных показателей: статический и
динамический аспекты» (Смуров, Полищук, 1989; см. в списке основных
публикаций).В весеннем семестре 2006 г. мною был прочитан курс лекций по общей
экологии для студентов кафедры биофизики (совместно с Д.В. Карелиным).
Специально для этого курса были разработаны контрольные задания (каждое задание
строилось на базе трехчасовой лекции и предлагалось в начале следующей лекции;
на его выполнение отводилось примерно полчаса).
Основные
научные публикации (в обратном хронологическом порядке)
Полищук Л.В., Файферберг
Я. Динамика массы тела дафний в свете теории жизненных стратегий: Анализ с
использованием метода вкладов // Журн. общ. биол. 2006. Т. 67. № 1. С. 23-36.
Об этой и предшествующей работе (Polishchuk, Vijverberg, 2005) см.: А. Марков. Размножаться лучше на сытый
желудок // Сайт «Элементы большой науки» http://elementy.ru/news/430179
Polishchuk L.V., Vijverberg
J. Contribution analysis of body mass
dynamics in Daphnia // Oecologia. 2005. V. 144. No. 2. P. 268-277.
Полищук Л.В. Скорость
размножения и угроза вымирания вида // Природа. 2003. № 7. С. 12-21. Об этой и
других работах этого цикла см.: А.М. Гиляров. Перспектива попадания вида в
Красную книгу предсказуема! // Природа. 2003. № 3. С. 78. http://vivovoco.rsl.ru/VV/NEWS/PRIRODA/2003/PR_03_03.HTM
Polishchuk L.V. Conservation priorities for Russian
mammals // Science. 2002. V. 297. P. 1123. Об этой работе см. J. Randerson. Glamorous animals get all the
cash // New Scientist. 2002. No. 2357. P. 9. http://www.newscientist.com/article/mg17523571.200.html
Polishchuk L.V. A fecundity to mortality ratio,
population size variability and the chance for a mammal species to be listed on
the Red List // Журн. общ. биол. 2002. Т. 63. № 2. С. 99-111.
Полищук Л.В., Цейтлин В.Б.
Масса тела, плотность популяции и число потомков у млекопитающих // Журн. общ.
биол. 2001.
Т. 62. № 1. С. 3-24.
Polishchuk L.V. Contribution analysis of
disturbance-caused changes in phytoplankton diversity // Ecology. 1999. V. 80.
No. 2. P. 721-725.
Mnatsakanova E.A., Polishchuk L.V. Role of
parthenogenetic natality and emergence from diapausing eggs in the dynamics of
some rotifer populations // Hydrobiologia. 1996. V. 320. P. 169-178.
Polishchuk L.V. Direct positive effect of invertebrate
predators on birth rate in Daphnia
studied with a new method of birth rate analysis // Limnol. Oceanogr. 1995. V.
40. No. 3. P. 483-489.
Полищук Л.В.
Избирательная элиминация и положительный эффект планктонных хищников на
рождаемость жертв // Докл. Академии наук. 1995. Т. 340. № 3. С. 437-439.
Polishchuk L.V. Population analysis and production
approach: Two trends in aquatic ecology // Russian J. Aquat. Ecol. 1992. V. 1.
No. 1. P.
3-8.
Полищук Л.В.
Популяционный анализ и продукционная гидробиология // Труды IV Поволжской конференции «Проблемы охраны вод и рыбных
ресурсов». Т. 1. Казань: Изд-во Казанского ун-та, 1991. С. 44-55.
Смуров А.В., Полищук Л.В.
Количественные методы оценки основных популяционных показателей: статический и
динамический аспекты. М.: Изд-во МГУ, 1989. 208 с.
Сахарова М.И., Полищук
Л.В. Опыт изучения динамики скорости роста планктонных животных (на примере Daphnia galeata из Можайского
водохранилища). 2. Когортные кривые роста // Зоол. журн. 1989. Т. 68. № 2. С.
178-187.
Полищук Л.В., Сахарова М.И.
Опыт изучения динамики скорости роста планктонных животных (на примере Daphnia galeata из Можайского водохранилища). 1. Факт динамики // Зоол. журн. 1988. Т. 67.
№ 12. С. 1812-1821.
Полищук Л.В., Сахарова
М.И. Относительное значение разных факторов в динамике рождаемости планктонных
животных // Докл. АН СССР. 1987. Т. 296. № 1. С. 253-256.
Полищук Л.В. Динамические
характеристики популяций планктонных животных. М.: Наука, 1986. 128 с.
Polishchuk L.V., Ghilarov A.M.. Comparison of two
approaches used to calculate zooplankton mortality // Limnol. Oceanogr. 1981.
V. 26. No. 6. P. 1162-1168.
Полищук Л.В., Романовский
Ю.Э. Теоретический подход к расчету продукции водных животных // Журн. общ.
биол. 1980. Т. 41. № 5. С. 645-654.
Полищук Л.В.
Сравнительная оценка различных методов определения рождаемости и смертности
планктонных животных // Журн. общ. биол. 1980. Т. 41. № 1. С. 125-137.